
���˾���[1]
ǰ��
������Ϊ��ˮ�����߲�Ϊ�������㷺������ֲ����֯�е�������ȩ��Ϊ����ĸ��϶����ࡣ�����Ͽɷ�Ϊԭ�������������ᣨͨ����Ϊ�������������ᡣ�����������1��ʾ��
��1�����ķ���
|
������Ĺ�����Ҳ��Ϊ�����ʣ� |
�����Թ�������ԭ���� |
||
|
�����Թ��� |
|
|
|
|
�ͼ��������� |
|||
|
������ |
|
||
������Ϊֲ��ϸ������ϸ���ڼ���в㣨����㣩����Ҫ�ɷֵ�ͬʱ��Ҳ��Ϊ���ʳɷִ�����ϸ���ڵ��ڲ�������ά�ء�����ά�ؼ������ʵȳɷֹ�ͬά��ϸ���ڵĹ��ܡ�����ϸ���ڵ�������ѧ���ʣ�������ķ������廯ѧ�ṹ�����Ӽ估������ϸ���ڳɷֵ�������йأ������������ϸ�����ڵķֲ��������йء���ϸ�������İ�����У�������ϸ��ճ�������ã��Ӷ�ʹ��֯�����ʵ��ļ���ԡ����ԡ������Եȵ���ѧ���ʡ����⣬�������ά�غͰ���ά�أ��������н�ǿ�ı�ʪ���ܣ�Ҳ��ϸ��������Ҫ�ı�ʪ�ɷ֡�
���������������º���ˮ���Ƿ�ʱ�����γ�����״���ʣ�����Ǵ�ˮ������ȡ�ģ���������ϣ�����ΪPectos
��һ���棬�����ڹ�ʵ�ij��켰ֲ����������ϻ�������ø�Թ�����Ӱ�죬�����ﻯѧ�Ҽ�ø��ѧ�ҵĹ�ע֮�£������ֽ�ø���о���ʼ������Ŀ���������ڣ����ܹ�ע��Ӧ����ֲ��ϸ���ں��������õ��Ĺ�����ʽ����ø������ø��2��~4�������⣬�����ƵĹ�ϵ�ϴ��ȼ�����߲��еĹ����ı仯�нϴ��Ӱ�졣���ң�������©�Ļ��й�����Ϊʳ����ά��Ч�á��ڴˣ����Ľ��Ԣٹ����Ļ�ѧ���ʡ��ڹ��������������۹����ķֽ�ø���ܹ��������ơ�����Ϊʳ����ά�Ĺ��������в�����
1.�����Ļ�ѧ����
��������������һֱ����Ϊ�dz����ṹ�ľ۰�����ȩ��ļ��������������ǺͰ����DZ��ӹ�����Ʒ�еķ��֣�֤���������ɰ������ǺͰ����ǵȹ��ɵĸ��϶����ࡣ���ң������е��������������������¿ɷֽ�ΪС���ӣ��Լ���ø���·ֽ�̶Ƚϵ͵�֤�����������ǵ���������5�������⣬ͨ��DEAE��ά��ɫ�����ɽ�ֻ����������ɵĶ����뺬����ȩ�����ڵ����Զ��ǽ������֣���һ��֤���ڹ�����Ʒ����������Ĵ���6��~8����
1.1 �����Ĵ���

ͼ1ϸ���ڵĹ���ģ��9��
��2 ����Ҷֲ���˫��Ҷֲ���ϸ���ڳɷ�10����Albersheim��9���ıȽ�
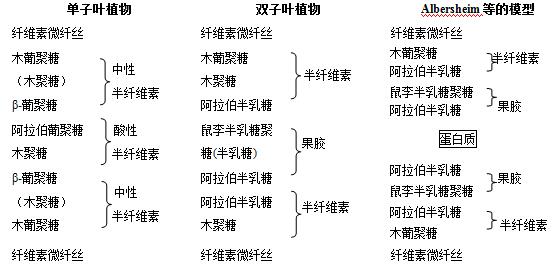
���еĹ��ɳɷּ�������״̬���б�ʾ�����ǣ�֮��������������ѧ�ķ��������ķ�չ�����˸�������Ϣ��ϸ����ģ��������顣��2��ʾΪ�����10�������ϸ���ڹ��ɳɷ���Albersheim�ıȽϡ��ڵ���Ҷֲ���˫��Ҷֲ���У�ϸ���ڵĹ��ɳɷ����Բ�ͬ�������Ԧ�-1,4-�Ͼ���Ϊ�����ṹ����������ά����˿����Ե���Ҷֲ���к������Զ��Ǽ�����������˫��Ҷֲ�������д�������������Ŀǰ��״Ϊ����ϸ�����еĶ�������Ӽ�Ľ�Ϸ�ʽ���Լ����ɷִ��ڵķ�ʽ�˽�IJ���̫ȫ�档
1.2 �����Ĺ����Ǻ�����
��ֲ����֯����ȡ�Ĺ���Ϊ������ȩ��Ϊ����Ķ����࣬�ж��������ǣ��������ڰ�����ȩ��������Ϲ����ϣ��������ǡ������ǡ�ľ�ǵ��ڲ����Ϲ����ϡ��Դӹ�ʵ����ȡ�Ĺ��������۹����Ĺ����ǹ۲��֪����Ϊ�����ǵİ�����ȩ��ĺ���Ϊ56%~85%���������Ǻ���Ϊ4%~14%12�����ܵ��������������ǺͰ����ǵĵĺ����ϸߣ���Ȼ�����ǵĺ������٣������й������к��С���ͬ��Դ�Ĺ��������һ���Ĺ����Ǻ��������ǿ���ȷ�ϵ��������Ǻ����ϸߵĹ����У������ǵĺ����ϵ͡����⣬������ȩ��Ĺ��������༰�����Թ������������dz���Ҫ�����á���ʵ�й��������۹����м������������3��ʾ��Ϊ���������������Ƴɼ����������ϵ͵Ĺ�������Ȼ��ͨ���ᡢ�ø����������4�ַ�������ͨ�����������Ѽ���ĵͼ����������У�������ȩ���C6�������Ȼ��������������������۾��ƹ����еİ���Ϊ5.5%��
��3�����м������ĺ�����%��
|
������Դ |
������* |
|
1�������� ���ָ� ���� ��� ���ݽ��� �Ľ� ���� ��� |
11.4 13.8 13.6 13.1 12.1 13.5 12.7 |
|
2��������ʵ�� ��
ƻ�� ף ӣ�� ���� ���� ���� ����� ÷�� |
12.9 11.8
10.1 9.2 11.6 11.7 12.6 13.1 9.4 |
|
3�����۹��� δ�������� ����NF ���������� �ͼ��������� ���ƹ��� ����NF ���������� �ͼ��������� |
12.1 11.7 6.3
12.8 11.8 6.1 |
*��ˮ������ȩ��base
1.3 �����ķ������ͷ��ӳ���
�����ķ������ͷ��ӳ���Ϊ��������ѧ���ʵ���Ҫ�������ӡ�ʹ�ù�ɢ��1Ũ�ȷ��ⶨ��ʵ������ƽ�������������Ϊ14~44��104��Conzalez-Chariere��14�������Ĺ����������У���ŷ�ΧΪ1~44��104���ֵķ�����Ϊ3~30��104�����⣬��ʾ���ӳ��ȵĹ��뾶Ϊ180~580Å��
ͨ�����뾶RG�ͷ�����MW�Ĺ�ϵ
��ĿǰΪֹ����Ȼ���ܳ���˽�����Ļ�ѧ�ṹ�����������Ĺ��죬�������Ȼ���������������ηֲ���������������С����⣬����Щ������̽�����Խ��ǽ����о����⡣
2. ������������
����������ͨ��״̬��Ϊ�����Һ����2���ɢϵͳ����״���ӣ����ࣩ�ĽӺ����������ͷ������ϱ�֯����״�ṹ���ڶ����Ĥ�ĸ������γ���ǻ������Щ��ǻ�������СҺ�Σ�Һ�ࣩ�������γɷ���������塣
ͼ2��ʾΪ���ǵ��������ṹģʽͼ������Ϊ����������ɵĹ���������

2.1 ����������������
��ͼ3��ʾ����������ȫ�����Ĺ����ļ���������ӦΪ16.32%����ͨ������½�7%���ϳ�ΪHM������7%���³�ΪLM��������Ȼ���ֹ�����ˮ��Һ��һ�������¿��γ����������������Ļ���ȴ����ͬ��
HM����-��-��-ˮ��ϵ�γɵ���������Ϊ�ǹ��ۼ���ϣ������ϣ��������������pH����ʱ���Ȼ����룬�Ƿ־��б�ʪ���ã���ͬ�ȶ�����������������������״���Ӽ�ͨ����״�Ӻ϶��γɵ���״�ṹ������ͨ�������ԵIJ��������Ϻ��γɵģ�Reese����17�������������Ϊ�Ӻ��������״�ص������и��ӵĻ�ѧ�ṹ��������δ�ɷ��˽�ʽ��ѶԹ���������������������ϸ˵����ͼ4��ʾΪ��Reese����16������İ����������ڵĶ����������ϵ�����������ģʽͼ��


���⣬LM����-�������-ˮ��ϵ�γɵ����������ں��и����ӣ�Ca2+���ȵ�2�۽��������ӣ��������ڵ���״���������ϵ������Ȼ������Žӣ��ڽӺ������γ���������ͼ5a��������Reese����16,17����Thibault18)�����ͼ5b��ʾ�ġ�����ģ�͡���
�Թ����������з��࣬���4��ʾ��LM����ʱ�������ӽ�ϼ���λ������ֽ�Ϸ�ʽ���ڡ�
��4 ���������ķ���
|
�������� |
��Ϸ�ʽ |
�Ӻ����� |
���� |
|
���������� ��HM�� |
�ǹ��ý�� �������ϣ� |
�������Ӽ�Ķ��������Ž� |
��������Ƥ���� |
|
�ͼ��������� ��LM�� |
���ӽ�� ��λ�Ӻ� |
�����������е��������������ӵ��µ������Ž� ���й¶Ե��ӶԵĹ��������������ӵ���λ��� |
ţ�̆�� ���� �����ⶳ |
2.2 ��ͬ���Ʒ����µ�LM������������������
ʹ�ð��������Ѽ���ALM���Ĺ�������Aspergillus japonicas�з�����Ĺ�����ø�Ѽ���ELM���Ĺ��������Ѽ���CLM���Ĺ���������Ca2+��Ϻ�ķ����������ӳ��Ƚ����о������Ϊ���з�������19��20��������CLM���������ALM����������Ca2+�����ӵ��·���������ӣ������ӳ��ȱ�С�����ӳ��ֳ�����״̬�����ң�ELM������ǰ���߲�ͬ����ȻҲ������Ca2+����������������ȴ�dz�С��Ishill����21���Ʋ�����ELM������Ca2+�������Դ���ALM������������γɽ�Ӳ��������
Padival����22������ģ����ᡢ�������Ƽ��������Ѽ���LM����������Ca2+���������������ԵĹ�ϵ��ͼ6��ʾ�����Ʒ�����ͬLM����Ҳ����ͬ������֮�������ŵIJ�ͬ���¹�������������������ͬ��

3. �����ֽ�ø
1960����ǰ�����ڹ�����ø�ֽ⣬����Ϊ�Ǽ�ˮ�ֽ�ø��ӵ����á�
��Neukom����23����ָ�������ڼ�����Һ�в��ȶ��������Ħ�-1,4��ϣ�ͨ���������ƽ��зֽ⣻��������������Һ�У�������Ҳ�ᷢ��ͬ���ķֽⷴӦ24��������ͨ��֮����о�25��~27�������ڹ����ķֽ⣬�м�ˮ�ֽ�ø�������ø2��ø������á������ֽ�ø�ķ������5��ʾ��
���⣬������ø�;۰�����ȩ��ø���Լ�������ʽ����ø�Թ����ķֽ������ͼ7��ʾ��
��������Ҷ�ϸ���ںϵĻ���Խ��Խ��ע��Ϊ����ϸ���ںϣ�������Ҫ������ϸ����ϸ���ڷֽ⣬ʹϸ��������¶״̬����������ʹԭ�����嵥�롣ͨ��ø��ʹԭ�����嵥��Ĺ�����ͼ84����ʾ�����ȣ�����Ҷ�Ĺ����ֽ�ʹ����ϸ���ڵ�ϸ�����룬Ȼ������ϸ����ϸ���ڷֽ⡣����Ҳ�н�����ø�����һ��һ�����ԭ������ķ�������Nagata����ʹ�õ�ԭ��������ø�����ø����Ϊ���н�����ǿ���ӵ�������ø�࣬���ܵ�ֲ��ϸ����Һ�о��ߵĹ�ע��

4. ����������
���������ƹ����еı仯���ܵ�ʳƷ�ʵص����ң���ʳ���ζ��������ϴ��Ӱ�졣���ڼ������߲˵Ĺ����ı仯�����Bartolome����28����ʹ���������й���ϸ�о�����������60~70���½���ǰ����������������������������Ϊ50������ʱ������ø�����л��ԣ��¶ȵ���50������ʱϸ��Ĥ�ܵ����ˣ�ϸ���ڵ���ʣ��ر���K+���ܳ���������ϸ���ڣ�ʹϸ����ø�Ĺ�����ø�����ˣ��������ļ����������������Ӽ��������Դ�ϸ��������Mg2+��Ca2+�����Žӣ�ʹ��֯Ӳ���������Ǽ���Ҳ����������
���⣬�������29����ʹ�����������ܲ�����ݼ���˻������ӡ����ѣ���60��Ԥ���ȵ�Ӳ����������о������õ���Bartolome����28����ͬ�Ľ����
���߲���������Һ�м������������������ڦ�-���루��ʽ���������¹������������϶��ѣ���ź��ţ��Ӵ���ʱ���������Ӳһ��Ļ�ʳ��ʱ���нϺõĿڸУ����������������������£���ʽ����ø���ܴٽ������ķֽ⡣
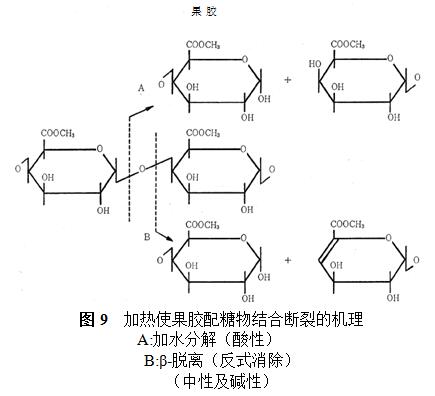
���ٵ���29�������������������Һ�У�pH7.1ʱ����Ѹ�ٱ���ʽ����ø�ֽ⣬pH6.3ʱ�����ֽ⣬pH4.1ʱ��û�з�Ӧ�������ǽ�ø��������ų����⣬������Һ�ڼ���ʱ��ǿ������Һͨ����ˮ�ֽ�ʹ�������϶��ѣ����Լ�������Һͨ����ʽ����ʹ�������϶��ѣ����û���������ͬ��������û�����ͼ9��ʾ��
5. ��Ϊʳ����ά�Ĺ���
ʳ����ά�Ķ���Ϊ�������ᱻ��������ø�������ġ�ʳ�����������Գɷֵ��ܳơ�31����ʳ����ά�ķ������Ҫ�ɷ����6��ʾ32��~34����
��6 ʳ����ά�ķ������Ҫ�ɷ�33��~35��
|
��Դ |
���� |
��Ҫ�ɷ� |
|
ϸ���ڵĽṹ���� |
���� |
��-D-�Ͼ��� |
|
����ά�� ������ά�ض����ࣩ |
ľ���� ��¶�� ������ |
|
|
�����������ԣ� |
�����Ǿ��� |
|
|
ľ���� |
������̼���������� |
|
|
�Ƕ��� |
�۰��������� |
|
|
�ǽṹ���� ����Ȼ���ʼ������ |
������ˮ���ԣ� |
�����Ǿ��� |
|
ֲ�コ |
����ȩ������ |
|
|
ճ���� |
�����Ǹ�¶�� ���Ǹ�¶�� |
|
|
������� |
����-���������� |
|
|
��ѧ���ζ��� |
�ӹ�����CMC |
����Ϊϸ���ڼ��в�Ĺ������ʣ�����ȡ���Ĺ���Ҳ����ʳ����ά�Ĺ�Ч��ͼ10��ʾΪ��������ģʽ�Ĺ��졢�����Լ���ʳ����ά����õij���32����ʳ����ά���г�������ʾ�Ĺ�Ч��
���������ĸ��ƺ��ȵ��ط��ڵ���ԼЧ����ͼ10��A��B��C��
����������ݣ�Jenkins����35��36��ʹ�ù϶��������������о������϶���������������У������������ڹ����У���������ʳ�ã�ʳ�ú�30~90�����ڣ�Ѫ�ǵ������ܵ��������ơ����⣬ʳ�ú��ȵ���ˮƽ������Ҳ�ܵ��������ơ�
�ڵ��̴���л��������Ч����ͼ10��D��E��F��G��
���ڵ��̴���л��������Ч�����н϶౨������1961�꣬Keys����37��1����ȡ15g��������ʹѪ�嵨�̴����͵����á�֮��Palmer����38��1����ȡ6~10g�������Լ�Durrington����39��1��12g��Jenkins����40��1��36gʱ��Ҳͬ����ʹѪ�嵨�̴����͡�
��ʳ�����к����ʵĶ����������ã�ͼ10��D��E��
�ܴ������ʵ�����Ч����ͼ10��F��G��
��732����ʾΪ������4��ʳ����ά�Ĺ�Ч����ͼ10����������A~G��λ�Ķ�Ӧ�ܽᡣ
���ϣ���һ��ʳ����ά��Ϊ���彡���ı���ʳƷ�ɷֵ�ԭ����в�����������������ˮ���Ի��Dz����ԣ�������Ҫʳ����ά�е�1�֡�

��7 ��������ʳ����ά����õij�����Ч��32��
|
��������λa�� |
ʳ����ά������ |
ʳ����ά��������� |
|
A |
���Ӿ����������ӱ����� |
SDFb����IDFc�� |
|
B |
��ˮ�����͡����������У����ƹ�����ȡ |
SDF��IDF |
|
C |
�ӳ�θ������ʱ�䣬���������� ��Լ�ȵ��صķ��� |
SDF��IDF������ |
|
D |
���Ƶ��̴������� ���͵��̴��������� ���ڵ��̴�Ũ�ȵ������� |
SDF����IDF |
|
E |
���͵�֭����������� |
SDF��������IDF |
|
D~E |
�ı�ʳ����ƶ��ٶȣ��ı����������صķ��ڣ����� �������������� ��ֹ�������ʵ��µ�Ӫ��������ȱ�� |
SDF��IDF |
|
F |
�ı䳦����ϸ��������֭���л�����̴���л �����°����ʵIJ��������� ���°����ʽ�ϻ���ϡ�� |
SDF��IDF |
|
G |
���ӵ��̴�����֭�ἰ��л�������й�� �����ű�������ű�˳�� |
SDF��IDF |